Seed plant introduction: Origin and evolution of the seed habit
The earliest seeds - Paleozoic era: Progymnosperms and seed ferns
Late Permian seed ferns: Gigantopterids and the origin of the angiosperms
Jurassic parc - Mesozoic era: Extinct gymnosperms and living ancient gymnosperms
Conifers - Life cycle of pine: Extant gymnosperms, the oldest trees
Seed Evolution Webpage 2 (this webpage)
Angiosperm seed evolution and species diversification - Darwin's abominable mystery
Seed phylogeny - Morphological and physiological trends in seed evolution
Structural seed types based on comparative internal morphology
Evolutionary trends of angiosperm seeds
|
 |
Angiosperm seed evolution and species diversification:
Darwin's abominable mystery
|
|
|
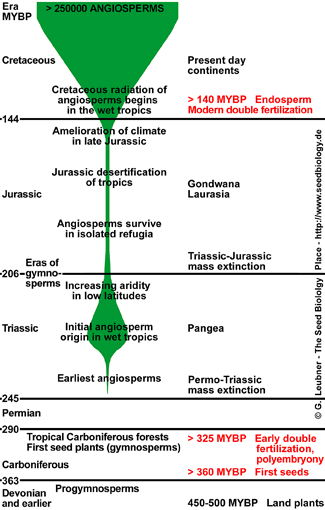
Angiosperms are the regarded as one of the greatest terrestrial radiations of recent geological times. This occurred in the Cretaceous era (see MYBP time scale figure): The major lineages originated 130-90 MYBP, following by a dramatic rise to ecological dominance 100-70 MYBP, and >250000 angiosperm species are known today. Charles Darwin described the rapid rise and early diversification within the angiosperms as 'an abdominable mystery'. This diversification is also manifested on the level of the seed, it's longevity and germination control is part of the species adaptation to environmental factors, e.g. to the season: temperature, light and water.
Among other reasons, the 'invention' the double fertilization that gives rise to seeds with triploid endosperm is proposed to be a major reason for
the evolutionary success of the angiosperms. Two hypotheses for the evolutionary origin of the endosperm are favored today (Friedman and Williams, 2004):
(1)
The endosperm is a sterilized homolog of an embryo (Sargant 1900, Friedman 1995)
(2) The endosperm is the sexualized homolog of a portion of the megagametophyte (Strasburger 1900, Coulter 1911)
Primitive angiosperms, e.g. Nymphaceae, have a 4-celled, 4-nucleate embryo sac and double fertilization gives rise to a diploid endosperm. Based on this and on additional findings, Friedman and Wiliams (2004) are in support for the hypothesis that this diploid endosperm originated from a supernumerary embryo (altruistic sibling embryo, hypothesis 1). Modular duplication resulted in the 7-celled, 8-nucleate embryo sac of most of today's angiosperms. In this case double fertilization gives rise to a triploid endosperm found in most angiosperms. Other researchers (e.g. Nowack et al., 2006) support hypothesis 2.
The endosperm is an embryo-nourishing tissue and is, depending on the species, parially or fully obliterated during seed development. However, most angiosperm species have retained an endosperm layer in the mature seed. In many of these cases, the endosperm not only functions as nourishing tissue, but is also involved in the control of seed germination in response to the environment and to developmental factors. Endosperm weakening is a prerequisite for the germination of many endospermic seeds including Solanaceae and Brassicaceae species.
Seed plant evolution hyperlinks
DT - Devonian Times by Dennis C. Murphy
UCMP - University of California Museum of Paleontology
Links for Paleobotanists - University Würzburg
Tree of Life Web Project (Tolweb)
Deep Time - Phylogenetic Tree of Angiosperms
Angiosperm Phylogeny Website (Stevens 2006, Missouri Botanical Garden)
Oleanane: Taylor et al., Paleobiology 32: 179-190 (2006)
Gigantopteroids: www.gigantopteroid.org (John M. Miller)
|
|
God to Noah
(after the great flood):
“As long as the earth endures,
seedtime and harvest,
cold and heat,
summer and winter,
day and night,
will never cease.”
Genesis 8,22 |
|
|
|
Seed phylogeny - Morphological and physiological trends in seed evolution
Seed biodiversity has attracted the attention of many researchers and is a hallmark of seed biology. This great diversity of morphological and physiological features have evolved to control germination and dormancy in response to different environments. The evolution of seed structure, germination and dormancy is summarized by the Tansley review by Bill Finch-Savage and Gerd Leubner (2006) and the references cited therein. The cited references include key literature to seed evolution like the book "Seeds" from Baskin and Baskin (1998), and the publications by Baskin and Baskin (2004), by Forbis et al. (2002), and by Nikolaeva (2004). The work on seed morphology is based on a publication by Martin (The comparative internal morphology of seeds. The American Midland Naturalist 36: 513-660, 1946).
The most obvious morphological difference in mature angiosperm seeds is their "embryo to seed" size ratios resulting from the extent to which the endosperm is obliterated during seed development by incorporating the nutrients into the storage cotyledons. Based on the internal morphology of 1287 mature seeds Martin (1946) defined the following seed types with distinct embryo to endosperm ratios:
Structural seed types based on comparative internal morphology
|
BASAL seed types (B1-B4) - basal division according to Martin (1946)
Embryos (black) usually relatively small, and restricted to the lower half of the seed, except in some of the lateral type B4. Seeds usually medium to large. Endosperm (orange) abundant and generally starchy
except in the rudimentary type B1. The seed types capitate B3 and lateral B4 are only found in monocots, wheras the seeds types rudimentary B1 and broad B2 are wider distributed.
|
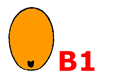 |
BASAL seed type B1- Embryo RUDIMENTARY
Abundant endosperm and tiny embryo. Embryos small, globular to oval-oblong, cotyledons usually rudimentary and obscure but sometimes they are evident. Seeds generally medium to large size. Examples: Magnolia spp. (Magnoliaceae, Magnoliales, Magnoliids, Basal Angiosperms); Anemone spp. and Trollius spp. (Ranunculaceae, Ranunculales, Basal Eudicots). |
 |
BASAL seed type B2 - Embryo BROAD
Embryo as wide as or wider than high, peripheral or nearly so. Examples: Nymphaea spp. (Nymphaceae, Nymphales, Basal Angiosperms). |
 |
BASAL seed type B3 - Embryo CAPITATE
Embryo expanded above into head-like form. Monocots only. Examples: Commelina spp. (Commelinaceae, Commelinales, Commelinids, Monocots). |
 |
BASAL seed type B4 - Embryo LATERAL
Embryo basal-lateral or lateral, inclined to expand in the plane of the periphery, small to half or rarely larger. Monocots only, typical for Poaceae seeds, e.g. cereal grains. Examples: Triticum spp. (Poaceae, Poales, Monocots). |
PHERIPHERAL seed type P - pheripheral division according to Martin (1946)
Embryos (black) ordinarily elongate and large, often curved, cotyledons narrow or expaneded. "Endosperm" (orange) conspicuously starchy, often central; "endosperm" ia actually perisperm in most cases.
|
 |
PHERIPHERAL seed type P - Embryo PERIPHERAL
Characteristics as described for the P-division. Examples: Beta vulgaris and Chenopodium album (Amaranthaceae, Caryophyllales, Caryophyllids). |
AXILE seed types (LA, MA, FA) - axile division according to Martin (1946)
Embryos (black) small to total, central (axile) straight, curved, coiled, bent or folded. Endosperm (orange) not strachy except in some monocot families of the LA-subtype. Well presented in Gymnosperms and Angiosperms. The axile division includes the linear, miniature and foliate subdivisions.
|
| LINEAR AXILE seed types (LA) - linear subdivision of the axile division according to Martin (1946) |
 |
LINEAR AXILE seed type LA - Embryo LINEAR AXILE
Embryo generally several times longer than broad, straight, curved or coiled, cotyledons not expanded. Seed size normally not minute. Examples: Pinus spp. (Pinaceae, Pinales, Gymnosperms); Allium spp. (Alliaceae, Liliales, Monocots); Iris spp. (Iridaceae, Asparagales, Monocots); Apium graveolens and Daucus carota (Apiaceae, Apiales, Asterids); Nicotiana spp. and Petunia hybrida (Solanaceae subfamily Cestroideae, Solanales, Asterids); Capsicum annuum and Lycopersicon esculentum (Solanaceae subfamily Solanoideae, Solanales, Asterids). |
MINIATURE AXILE seed types (MA) - miniature subdivision of the axile division according to Martin (1946) |
 |
MINIATURE AXILE seed types MA - Embryo MINIATURE AXILE
Seeds small to minute with embryos that are stocky or minute; seed coverings generally delicate and often cellular-reticulate. Endosperm not starchy. Within the MA-seed type subdivision Martin (1946) distinguished between dwarf or micro MA-seed types.
The evolution of the small seed size (e.g. dwarf seeds, MA) is described elsewhere, e.g. in Moles et al. (2005) and in Baskin and Baskin (2005) and Baskin and Baskin (2007). Small embryos in dwarf seeds may or may not be undifferentiated (not differentiated into organs) or underdeveloped (differentiated, but needs to grow). The assignment of seeds to a dormancy class requires that studies be done to determine if embryos grow inside the seed before germination can occur. Embryos of Drosera anglica are small, but fully developed, they don't grow in the seed prior to germination (seeds have PD). Embryos of Campanula americana are underdeveloped and need to grow prior to germination (seeds have MD).
DWARF (REMOVED AS SEED TYPE) seed size is 0.2 to 2 mm long. Embryos are varible in relative size, small to total, generally stocky, usually oval to elliptic or oblong. Cotyledons inclined to poorly developed. Examples: Drosera angelica (Droseraceae, Caryophyllales, Caryophyllids); Campanula americana (Campanulaceae, Asterales, Asterids); Begonia spp. (Begoniaceae, Cucubirtales, Rosids).
MICRO (REPLACED BY UNDIFFERENTIATED AS SEED TYPE) seed size is < 0.2 mm, i.e. minute-sized seeds. Embryo minute to total. Testa generally globular and consisting of relatively few cells (ca. 50-150).
Examples: Orchidaceae species.
REVISION OF THE MA-TYPE (2007): A revision of Martin's seed classification system, with particular reference to his dwarf-seed type was proposed by Baskin and Baskin (2007). According to this revision, the dwarf category has been removed and the micro category replaced by UNDIFFERENTIATED to reflect the state of the embryo in fresh seeds. The "dwarf seed type" per se is of limited value to seed biologists and these seeds are assigned to the other ten seed types irrespective of their size. Martin's conclusion that dwarf seeds are more advanced in evolution compared to medium or large seeds with rudimentary embryos. However, according to this revision, seed size is not longer a criterium for the assignment to a specific seed type.
The "micro seed type" is now replaced by a "undifferentiated seed type", indicating that the embryo in fresh seeds lacks organs. Most of these seeds are of micro-size, but there are also relatively large seeds whose embryos lack organs at the to time of maturity.
|
FOLIATE AXILE seed types (FA1-FA4) - foliate subdivision of the axile division according to Martin (1946)
Embryos (black) large, generally quarter to total, central rather than peripheral. Cotyledons expanded. Seeds generally medium to large. Endosperm (orange) generally not starchy. In mature FA2, FA3, and FA4 seed types the embryo occupies most of the seed; there is little or no endosperm, the nutrients are stored in the cotyledons.
|
 |
FOLIATE AXILE seed type FA1 - Embryo SPATULATE AXILE
Embryo spatulate axile (erect). Cotyledons thin to thick and slightly expanded to broad. Examples: Ephedra spp. (Ephedraceae, Gnetales, Gymnosperms); Fraxinus excelsior and Syringa spp. (Oleaceae, Lamiales, Asterids); Lactuca sativa and Helianthus annuus (Asteraceae, Asterales, Asterids); Vitis spp. (Vitaceae, Vitales, Rosids); Ricinus communis (Euphorbiaceae, Malphighiales, Rosids); Cucumis spp. (Cucurbitaceae, Cucurbitales, Rosids). |
 |
FOLIATE AXILE seed type FA2 - Embryo BENT AXILE
Embryo bent axile (spatulate but bent in jackknife form). Cotyledons generally thick. Examples: Rhus spp. (Anacardiaceae, Sapindales, Rosids); Trifolium spp., Melilotus alba, Glycine max and Pisum sativum (Fabaceae, Fabales, Rosids); Lepidium spp., Arabidopsis spp., and Brassica spp. (Brassicaceae, Brassicales, Rosids). |
 |
FOLIATE AXILE seed type FA3 - Embryo FOLDED AXILE
Embryo folded axile. Cotyledons usually thin, extensively expanded and folded in various ways. Examples: Most Acer spp. (Sapindaceae, Sapindales, Rosids); Geranium spp. and Pelargonium spp. (Geraniaceae, Geraniales, Rosids); Malva rotundifolia (Malvaceae, Malvales, Rosids). |
 |
FOLIATE AXILE seed type FA4 - Embryo INVESTING AXILE
Embryo investing axile (erect). Cotyledons thick, overlapping, encasing the somewhat dwarfed stalk for at least half its length. Endosperm wanting or limited. Examples: Fagus sylvatica (Fagaceae, Fagales, Rosids); Ceanothus spp. (Rhamnaceae, Rosales, Rosids). |
| |
 |
Evolutionary trends of angiosperm seeds
Martin (The comparative internal morphology of seeds. The American Midland Naturalist 36: 513-660, 1946) arranged the seed types in a seed phylogenetic tree (see below) and proposed evolutionary seed trends. This has been revised and extended by Forbis et al. (2002) and is presented in the Tansley review by Finch-Savage and Leubner-Metzger (2006). In summary, the following general evolutionary seed trends are obvious:
(1) In mature seeds of primitive angiosperms a small embryo is embedded in abundant endosperm tissue. Such seed types (e.g. B1) are prevailing among basal angiosperms.
(2) The general evolutionary trend within the higher angiosperms is via the LA seed type (embryo linear axile and developed, endosperm abundance medium to high) towards FA seed types (embryo foliate axile and developed, often storage cotyledons, endosperm abundance low or endosperm obliterated) with storage cotyledons. The LA seed-type is typical for many Asterids, e.g. the endospermic Solanceae seeds. Further embryo dominance and endosperm reduction leads via the FA1 seed type to the diverted seed types FA2, FA3 and FA4. The FA seed types are typical for many Rosids, e.g. Brassicaceae seeds with more or less no endosperm at maturity.
(3) In addition to these general seed trends there are clade-specific seed type differences ("exceptions"), e.g. within the basal angiosperms (Laurales) and the Asterids (Aquifoliales).
(4) A small embryo is also found in primitive gymnosperms and an increase in the E:S values is also evident within the gymnosperms. An increase in relative embryo size appears therefore to be a general evolutionary trend within the angiosperms and the gymnosperms.
These morphological trends in seed evolution are of utmost importance for seed physiology, especially for the evolution of seed dormancy.
The evolution of whole seed size (e.g. dwarf seeds, MA) is discussed elsewhere, e.g. in Baskin and Baskin (2005), Baskin and Baskin (2007), and in Moles et al. (2005).
|
|
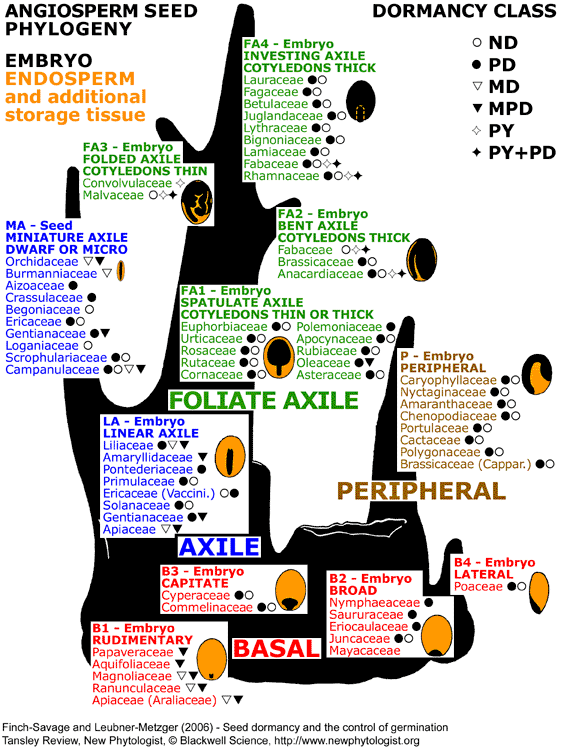 |
|
Seed phylogenetic tree based on the internal morphology of embryo and endosperm in mature angiosperm seeds.
Martin (1946) investigated the embryo (form, size, position) and endosperm (plus additional storage tissue) in 1287 species and proposed seed types (B1 to B4, LA, P, MA, FA1 to FA4). Seed types with abundant endosperm (orange) and a tiny embryo (black) are basal (B1, B2, B3, B4). In the more advanced endospermic LA-type seeds, the embryo is linear axile. From this developed the FA-type seeds (FA1, FA2, FA3, FA4) where the embryo is foliate axile and, depending on the subtype, differs in shape and occupies more or less the entire seed. Mature FA-type seeds have little or no endosperm, and the nutrients are stored in the cotyledons.
Seed dormancy classes are indicated next to each family name: non-dormancy (ND), physiological dormancy (PD), morphological dormancy (MD), morphophysiological dormancy (MPD), physical dormancy (PY), combinatorial are explained on the web page "seed dormancy".
Updated and modified from Martin (1946) based on work from Baskin and Baskin: 1998, 2004, 2005, and personal communication.
Figure published in the
Tansley review "Seed dormancy and the control of germination" by Finch-Savage and Leubner-Metzger (New Phytologist 171: 501-523, 2006).
A PDF file of this review is available for download from
either "The Seed Biology Place" (Finch-Savage and Leubner-Metzger, 2006)
or the New Phytologist Trust website at New Phytologist Trust - Tansley Reviews -
© Blackwell Science -
http://www.blackwell-science.com/
|
|
| |
|
|
|
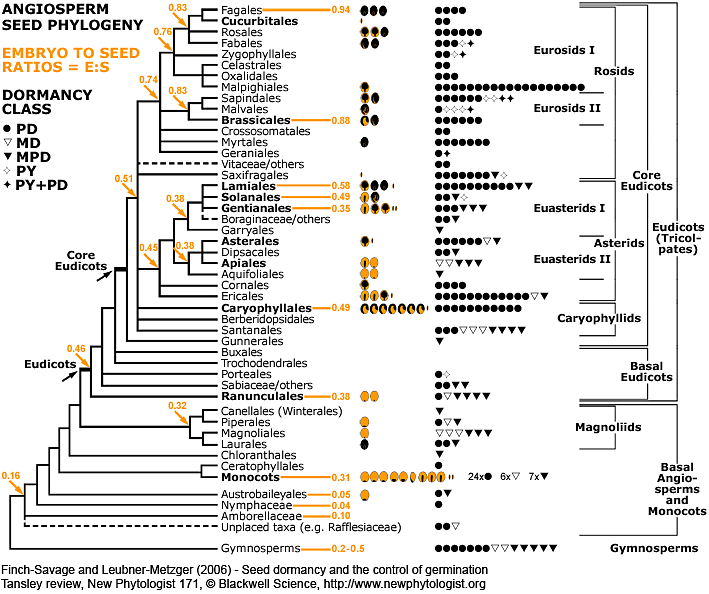 |
|
Angiosperm seed evolution depicted in a phylogenetic tree of the Angiosperm Phylogeny Group II (2003).
For each family, pictographs of the seed type and symbols for the dormancy class are placed next to the corresponding clade (see figure above for symbols and data source). The number of pictographs or symbols is equal to the number of families with the corresponding seed type or dormancy class, respectively. Numbers in the phylogenetic tree represent "embryo to seed" (E:S) ratios expressed as GSL (generalized least squares) values from Forbis et al. (2002, personal communication).
Phylogenetic tree modified from Judd et al. (2002); Angiosperm Phylogeny Group (2003); Soltis and Soltis (2003); Stevens (2006). Figure published in the
Tansley review "Seed dormancy and the control of germination" by Finch-Savage and Leubner-Metzger (New Phytologist 171: 501-523, 2006). A PDF file of this review is available for download from
either "The Seed Biology Place" (Finch-Savage and Leubner-Metzger, 2006)
or the New Phytologist Trust website at
New Phytologist Trust - Tansley Reviews -
© Blackwell Science -
http://www.blackwell-science.com/
|
|
| |
|
|
|
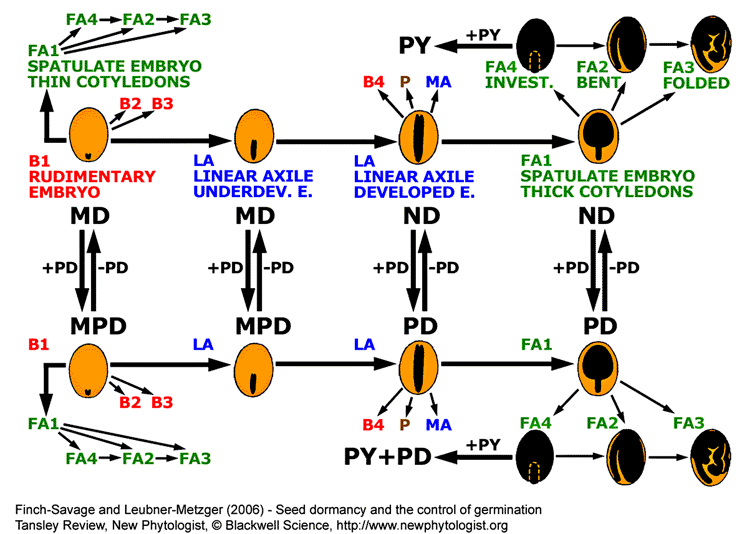 |
|
Summary of general trends in angiosperm seed evolution with respect to seed type and dormancy class.
Seeds with small embryo and abundant endosperm (B1 seed type) are primitive and evolution shows a general trend (indicated by arrows) towards mature seeds with little or no endosperm in which the embryo occupies most of the seed (through LA to FA seed types). Gain or loss of PD to each of these seed types (B1, LA and FA1) alters the dormancy class as indicated. Similar changes can also occur in the other seed types. Addition of PY to FA4-type ND or PD seeds is indicated as a side branch occurring in specialized species. This addition can be reversed and PY can also be added to other seed types (not shown). Seed types and dormancy classes are described in the figure above. This diagram is based on information given in figures 12.21 and 12.22 of Baskin and Baskin (1998). Figure published in the
Tansley review "Seed dormancy and the control of germination" by Finch-Savage and Leubner-Metzger (New Phytologist 171: 501-523, 2006). A PDF file of this review is available for download from
either "The Seed Biology Place" (Finch-Savage and Leubner-Metzger, 2006)
or the New Phytologist Trust website at New Phytologist Trust - Tansley Reviews -
© Blackwell Science -
http://www.blackwell-science.com/
|
|
|
|
| |
|
|
|
| Anigosperms (flowering plants) appeared in the Lower Cretaceous and came to dominate most terrestial ecosystems. |
|
|
|
| |
|
|
|
|
 |
| |
|
|
|

|
|
