|
|
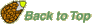
Seed germination: definition and reviews
- Seed germination: Germination of seeds is a complex physiological process triggered by imbibition of water after possible dormancy mechanisms have been released by appropriate triggers (see webpage "Seed Dormancy").Under favorable conditions rapid expansion growth of the embryo culminates in rupture of the covering layers and emergence of the radicle. Radicle emergence is considered as the completion of germination. The definition that a visible protrusion of radicle tip is the completion of germination is not only a definition issue of seed physiologists. This transition point is also characterized by the loss of dessication tolerance and this is a molecular checkpoint (in Arabidopsis regulated by ABI5), a developmental molecular switch from the germination program to the seedling program.
- Non-dormant seeds: A completely non-dormant seed has the capacity to germinate over the wides range of normal physical environmental factors possible for the genotype (Finch-Savage and Leubner-Metzger, 2006) Besides the basic requirement for water, oxygen and an appropriate temperature the seed may also be sensitive to light and/or nitrate. Germination commences with the uptake of water by imbibition of the dry seed, followed by embryo expansion. This usually culminates in rupture of the covering layers and emergence of the radicle, generally considered as the completion of germination. Radicle protrusion at the completion of seed germination therefore depends on embryo growth driven by water uptake.
- Cell elongation is necessary and is generally accepted to be sufficient for the completion of radicle protrusion, cell division is not essential. Thus, germination is a process and the completion of germination is an event; visible protrusion of the radicle tip through all covering structures is the typical criterion for the completion of seed germination.
- Uptake of water by a mature dry seed is triphasic (see webpage "Water Relations"), with a rapid initial uptake (phase I, imbibition) followed by a plateau phase (phase II, metabolic preparation for germination). Phase III is a further increase in water uptake which occurs directly after germination is completed. Phase-III water uptake causes hydraulic growth of the embryo and the emerged seedling. The plant hormone ABA inhibits phase III-water uptake.
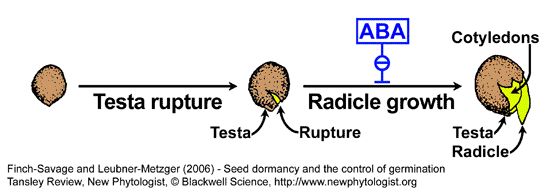
- In many species the covering tissues, e.g. endosperm and testa (seed coat), act as a physical barriers which must be overcome by the growth potential of the embryo if the seed is to complete its germination (Linkies et al., 2010). In coat-dormancy, the seeds are prevented from completing germination because the embryo is constrained by the covering layers. The micropylar endosperm is known since a long time as a constraining structure in members of the Asteraceae (e.g. lettuce) and Solanaceae (e.g. tomato, tobacco and Datura spp.). Endosperm weakening, at least limited cell wall hydrolysis, by the action of specific hydrolases seems to be needed for the completion of germination by endosperm rupture. In addition to the endosperm, the micropylar testa also confers a hindrance for radicle protrusion of of tobacco and tomato seeds.
- Arabidopsis thaliana and Lepidium sativum are Brassicaceae seeds with distinct testa rupture and endosperm rupture. Müller et al. (2006) demonstrated that endosperm weakening occurs prior to Lepidium sativum endosperm rupture and is controlled by the GA-ABA ratio. Linkies et al. (2009) demonstrated that ethylene promotes endosperm cap weakening of Lepidium sativum and endosperm rupture of the close Brassicaceae relatives Lepidium sativum and Arabidopsis thaliana. Ethylene counteracts the inhibitory action of abscisic acid (ABA) on these two processes.
- A comprehensive table of Arabidopsis homone mutants summarizes the altered phenotypes regarding germination and dormancy.
- Recommended reviews on seed germination:
"First off the mark: early seed germination" by Weitbrecht, Müller, Leubner-Metzger,
Darwin review: Journal of Experimental Botany in press (2011)
"Seed dormancy and the control of germination" by Finch-Savage and Leubner-Metzger,
Tansley Review, New Phytologist 171: 501-523 (2006)
"Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination" by Holdsworth, Bentsink, Soppe,
Tansley Review, New Phytologist 179: 33-54 (2008)
"Plant hormone interactions during seed dormancy release and germination" by Kucera, Cohn, Leubner-Metzger,
Seed Science Research 15: 281-307 (2005)
"Functions and regulation of ß-1,3-glucanase during seed germination, dormancy release and after-ripening" by Leubner-Metzger,
Seed Science Research 13: 17-34 (2003)
"Seed dormancy and germination" by Koornneef, Bentsink, and Hilhorst,
Curr Opin Plant Biol 5: 33-36 (2002) (download 1.3 MB PDF-file)
"Seed germination and dormancy" by Bewley,
Plant Cell 9: 1055-1066 (1997) (download 1.9 MB PDF-file)
|
One-step seed germination of Brassica and pea seeds: testa rupture and initial radicle elongation
- The endosperm is completele obliterated during the seed development of Brassica spp. (see figure below) or pea and the mature seeds of these species are therefore non-endospermic. Uptake of water by a seed is triphasic with a rapid initial uptake (phase I, i.e. imbibition) followed by a plateau phase (phase II). A further increase in water uptake (phase III) occurs only when germination is completed, as the embryo axes elongates and breaks through the testa. Thus, besides radicle elongation, testa rupture is the only visible landmark during Brassica spp. and pea seed germination.
- Abscisic acid (ABA) does not inhibit imbibition and testa rupture (see figure below), but ABA inhibits phase III water uptake and the transition from germination to postgermination growth (e.g. Schopfer and Plachy, 1984; Manz et al., 2005).
Brassica napus seed germination is one-step. The mature seeds of these species are without endosperm and so testa rupture plus initial radicle elongation result in the completion of germination. ABA does not inhibit testa rupture, but inhibits subsequent radicle growth (Schopfer & Plachy, 1984). Review: Finch-Savage and Leubner-Metzger (2006).
|
Two-step seed germination of Lepidium and Arabidopsis (Brassicaceae): testa and endosperm rupture
- For the Lepidium and Arabidopsis seed anatomy see the webpage "Seed Structure".
- Rupture of the testa (seed coat) and rupture of the endosperm are separate events in the germination of Lepidium and Arabidopsis seeds (see figures below).
Arabidopsis (Liu et al., Plant J 41:936-944, 2005) and Lepidium (Müller et al., 2006) exhibit a two-step germination, in which testa rupture and endosperm rupture are sequential events.
- Such two-step germination is widespread over the entire phylogenetic tree and has been described for many species, e.g. for Trollius (Ranunculaceae; Hepher and Roberts 1985), Chenopodium (Amaranthaceae; Karssen 1968; Karssen 1976), Nicotiana and Petunia (Cestroideae subfamily of the Solanaceae, Krock et al. 2002; Leubner-Metzger et al. 1995; Petruzzelli et al. 2003).
- We found that the plant hormone ABA inhibits endosperm rupture, but not testa rupture, of Arabidopsis and Lepidium (Müller et al., 2006). This inhibitory effect of ABA is counteracted by GA, supporting the view that endosperm rupture is under the control of an ABA-GA antagonism (Kucera et al. 2005).
- We found that ABA inhibits endosperm weakening of Lepidium, and this inhibitory effect is counteracted by GA (Müller et al., 2006). This supports the view that weakening of the micropylar endosperm occurs in Arabidopsis and Lepidium seeds (Brassicaceae, Rosid clade), is under ABA-GA control, and is functioning in controlling the germination of endospermic Brassicaceae seeds.
- We show that ethylene promotes endosperm cap weakening of Lepidium sativum and endosperm rupture of the close Brassicaceae relatives Lepidium sativum and Arabidopsis thaliana and that it counteracts the inhibitory action of abscisic acid (ABA) on these two processes (Linkies et al., 2009). Cross-species microarrays of the Lepidium micropylar endosperm cap and the radicle show that the ethylene-ABA antagonism involves both tissues and has the micropylar endosperm cap as a major target. Ethylene counteracts the ABA-induced inhibition without affecting seed ABA levels. The Arabidopsis loss-of-function mutants ACC oxidase2 (aco2; ethylene biosynthesis) and constitutive triple response1 (ctr1; ethylene signaling) are impaired in the 1-aminocyclopropane-1-carboxylic acid (ACC)-mediated reversion of the ABA-induced inhibition of seed germination. Ethylene production by the ACC oxidase orthologs Lepidium ACO2 and Arabidopsis ACO2 appears to be a key regulatory step. Endosperm cap weakening and rupture are promoted by ethylene and inhibited by ABA to regulate germination in a process conserved across the Brassicaceae (Linkies et al., 2009).
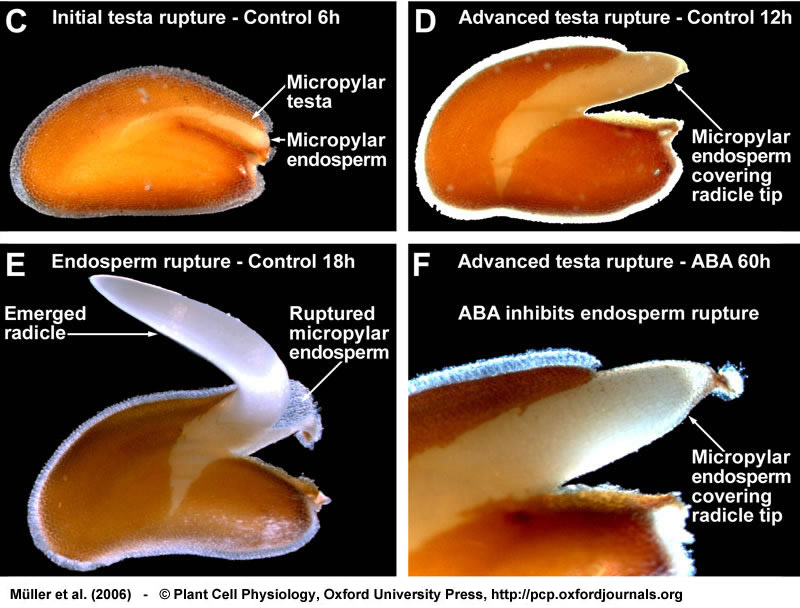
Two-step germination of Lepidium sativum. (C-F) During the two-step germination of Lepidium testa rupture (C,D) is followed by endosperm rupture, which occurs after 16 h under control conditions (E). Due to the microphotographic settings the transparent outer mucilage layer is not visible. (F) ABA specifically inhibits endosperm rupture, the radicle remains covered by the micropylar endosperm even after 60 h incubation in the presence of ABA. (G) Drawing of a mature Arabidopsis seed; the seed anatomy that is very similar to that of Lepidium. (H-J) Arabidopsis seeds also germinate with testa rupture (H) preceding endosperm rupture (I). Also during the two-step germination process of Arabidopsis, ABA specifically inhibits endosperm rupture (J). Seeds were incubated in continuous light without (control) or with 10 µM ABA added to the medium. From Müller et al., (2006).
|
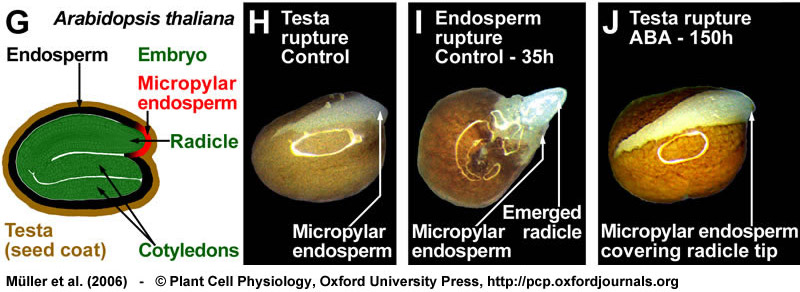
Two-step germination of Arabidopsis thaliana. (G) Drawing of a mature Arabidopsis seed; the seed anatomy that is very similar to that of Lepidium. (H-J) Arabidopsis seeds also germinate with testa rupture (H) preceding endosperm rupture (I). Also during the two-step germination process of Arabidopsis, ABA specifically inhibits endosperm rupture (J). Seeds were incubated in continuous light without (control) or with 10 µM ABA added to the medium. From Müller et al., (2006).
A comprehensive table of Arabidopsis hormone mutants summarizes the altered phenotypes regarding germination and dormancy.
|
|
|
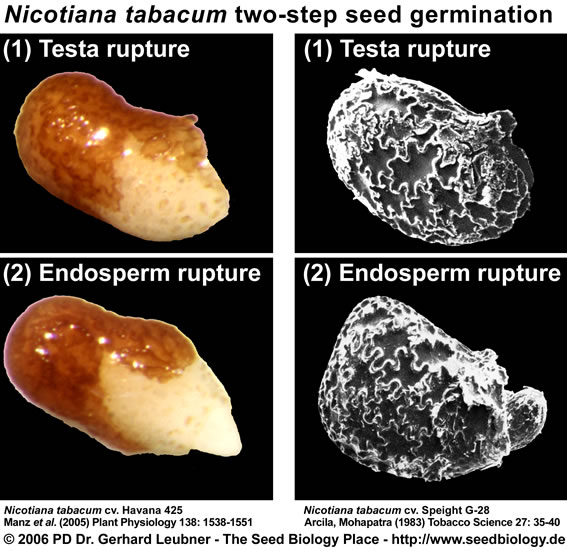
Two-step seed germination of Nicotiana spp. (Solanaceae): testa and endosperm rupture
- For the tobacco seed anatomy see the webpage "Seed Structure".
- Rupture of the testa (seed coat) and rupture of the endosperm are separate events in the germination of tobacco seeds and there is strong evidence that both, testa rupture and endosperm rupture are the limiting factors in the germination of these seeds (reviewed by Leubner-Metzger 2003).
|
- Electron microscopic studies support the view that the endospermic hole of the germinated tobacco seed, which is always at the micropylar end, is formed by "dissolution" rather than by "pushing" action (Arcila and Mohapatra, Tobacco Science 27: 35-40, 1983). Endosperm weakening is a pre-requisite for the completion of germination of many Solanaceous seeds.
- In photodormant varieties of tobacco that do not germinate in darkness, both the testa rupture and endosperm remain intact in the photodormant seeds. However, when the testa and endosperm are mechanically removed, there is radicle growth even in the absence of light. This shows that in tobacco coat-imposed dormancy is more important than embryo dormancy.
- Treatment of tobacco seeds with 10 µM abscisic acid (ABA) greatly delays endosperm rupture, but not testa rupture, and results in the formation of a novel structure consisting of the enlarging radicle with a sheath of greatly elongated endosperm tissue.
|
|
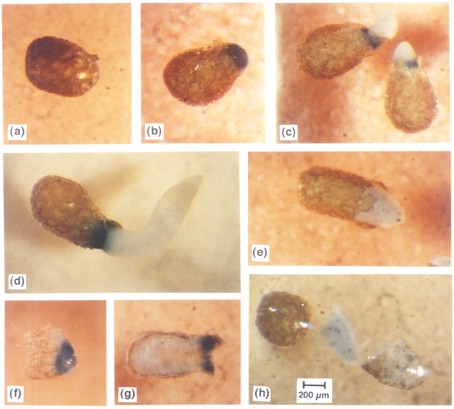
Figure 2:
Leubner-Metzger et al. (1995)
Testa rupture (b) and endosperm rupture (c) are separate events during the germination of tobacco seeds.
ABA delays endosperm rupture, but not testa rupture (e and h). |
Figure 2: Stages in the germination of tobacco seed homozygous for the GLB-GUS transgene. Seeds were germinated in continuous light at 24 °C with 10 µM ABA (ABA) and without ABA (control) added to the medium. At the times indicated after the start of imbibition, seeds were stained for GUS activity. The blue staining is indicative of transcriptional activity of the class I ß-1,3-glucanase B promoter. (a) Stage I (control, 3 h): intact seed prior to seed-coat rupture. (b) Stage II (control, 60 h): seed with ruptured seed coat and protruding endosperm. (c) Stage III (control, 72 h): seeds with ruptured endosperm showing GUS staining at the rupture site and emerging radicles, which do not stain for GUS. (d) Stage III (control, 96 h): seed with ruptured endosperm and elongating radicle. GUS staining is localized in a collar of endosperm tissue at the site of radicle penetration. (e) Stage II (ABA, 144 h): ABA treatment markedly delays endosperm rupture and results in a novel structure consisting of the enlarging radicle completely enclosed in a sheath of intact endosperm, which does not stain for GUS. (f) Stage II (control, 60 h): endosperm dissected prior to rupture showing GUS stain localized in the micropylar region. (g) Stage III (control, 96 h): endosperm dissected after rupture showing that the radicle penetrates the region which stains for GUS. (h) Stage II (ABA, 144 h): a seed arrested in stage II by ABA treatment dissected to show that the elongated radicle is enclosed in a sheath of endosperm. Magnification: 40X. |
|
| |
|
 |
| |
|
|
|

|
|


